nature reviews neurology|精神分裂症:从神经化学到环路、症状和治疗
原创nature reviews neurology|精神分裂症:从神经化学到环路、症状和治疗
原创
摘要:精神分裂症是全球致残的主要原因。目前的药物治疗主要使用一种机制-多巴胺D2受体阻断,但结果往往显示出有限的疗效和耐受性差。这些限制突出了需要更好地了解疾病的病因,以帮助发展替代治疗方法。在这里,我们回顾了最新的荟萃分析和其他关于前驱、首发和慢性精神分裂症的神经生物学研究结果,以及它们与精神病症状的联系,重点是来自精神分裂症患者的影像学证据。这一证据表明,与健康个体相比,区域特异性神经递质改变,包括基底神经节谷氨酸和多巴胺含量较高。我们考虑皮质-丘脑-纹状体-中脑回路的功能障碍如何改变大脑信息处理,从而成为精神病症状的基础。最后,我们讨论了这些发现对开发新的、基于机制的治疗方法和精确医学对精神病症状、阴性症状和认知症状的影响。
1. 引言
精神分裂症是全球致残的主要原因,影响约1%的人,通常在成年早期发病。在做出诊断之前,症状至少需要出现一个月,尽管它们通常持续更长时间。精神分裂症还与认知障碍有关,这是表现的一个常见部分,大约三分之二的精神分裂症患者都有这种症状。阴性和认知症状通常在首次精神病发作前数月至数年出现。在大多数个体中,症状表现为慢性、复发缓解或持续病程,尽管约10%的个体在一次发作后恢复。
由于精神分裂症发病于成年早期、社会和职业障碍以及总体预期寿命缩短,因此会带来巨大的经济和社会成本。目前所有获得许可的精神分裂症药物都是多巴胺D2/3受体阻滞剂,这是主要的治疗方法,但对阴性症状和认知症状的疗效有限。此外,虽然在许多个体中有效减轻精神病症状,但15-30%的人出现治疗耐药性,许多人报告了与D2/3阻断相关的难以忍受的不良反应。这些问题突出了根据疾病的病因发展替代治疗方法的必要性。在这里,我们回顾了精神分裂症的病理生理学,重点关注体内神经化学改变和电路,可能是药物治疗的目标,然后考虑潜在的新兴治疗方法。
2. 精神分裂症中的多巴胺
多巴胺是一种单胺神经递质,在大脑中介导信号转导。大多数多巴胺能神经元胞体位于中脑,特别是在黑质和腹侧被盖区,并有密集的投射到纹状体,但也投射到额叶皮层和其他几个大脑区域。最初的尸检和神经影像学研究表明,与健康对照者相比,精神分裂症患者中脑中多巴胺标记物的水平发生了改变。然而,研究的主要焦点是纹状体,在50多个分子成像研究中已经显示了纹状体改变的证据。直到过去十年左右,人们还不清楚精神分裂症患者多巴胺能系统的具体分子和回路变化。分子成像技术的应用有助于解决这一知识差距。
迄今为止,已有超过20项分子成像研究对慢性或首发精神分裂症患者多巴胺能系统的突触前方面进行了研究。多巴胺是由其前体左旋多巴(L-DOPA)通过芳香酸脱羧酶合成的。放射性标记的左旋多巴可以与正电子发射断层扫描(PET)一起使用,以指示左旋多巴在体内的摄取和转化为多巴胺,作为多巴胺合成能力的衡量标准。另外,多巴胺释放能力是通过D2/3受体配体结合的变化来指示的,这些变化是在释放多巴胺的挑战之后,如安非他明或社会心理压力。使用这些技术的研究荟萃分析显示,与健康个体相比,精神分裂症患者纹状体多巴胺合成和释放能力更高,且效应量大 (图1)。
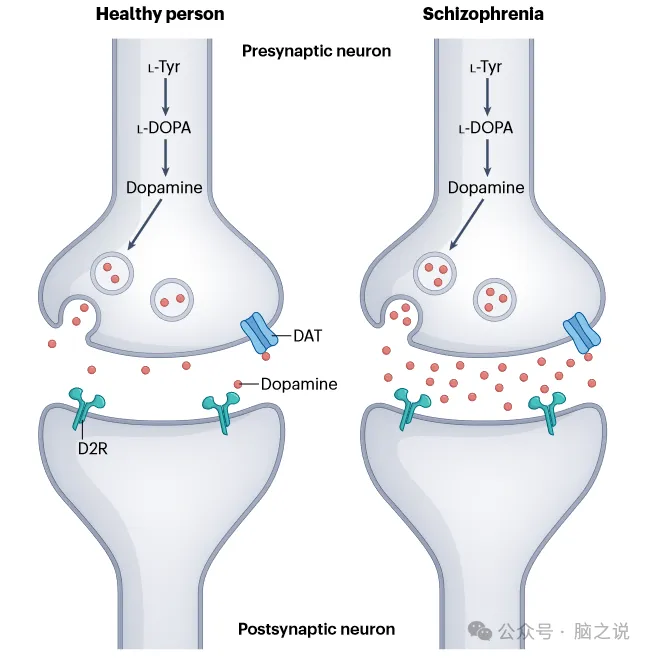
图1 健康个体和精神分裂症患者的多巴胺能突触。多巴胺的合成途径(红圈),显示多巴胺在囊泡中储存并释放到突触中,神经递质可以与多巴胺受体或转运体结合。在精神分裂症患者中,多巴胺的合成和释放能力以及突触多巴胺的水平高于健康人。
其他研究调查了精神分裂症高危人群的多巴胺能系统,因为他们被证实是高渗透性遗传风险因素的携带者,或者因为他们表现出前驱型的临床特征,称为临床高风险。与对精神分裂症患者的观察一致,这些个体也表现出相对于年龄和性别匹配的健康个体更高的纹状体多巴胺合成和释放能力。这些改变在随后继续发展为精神病的个体中最为明显,纹状体多巴胺合成能力的增强与更严重的亚临床症状直接相关。纹状体多巴胺合成能力的纵向增加也在前驱症状和首次精神病发作之间观察到。
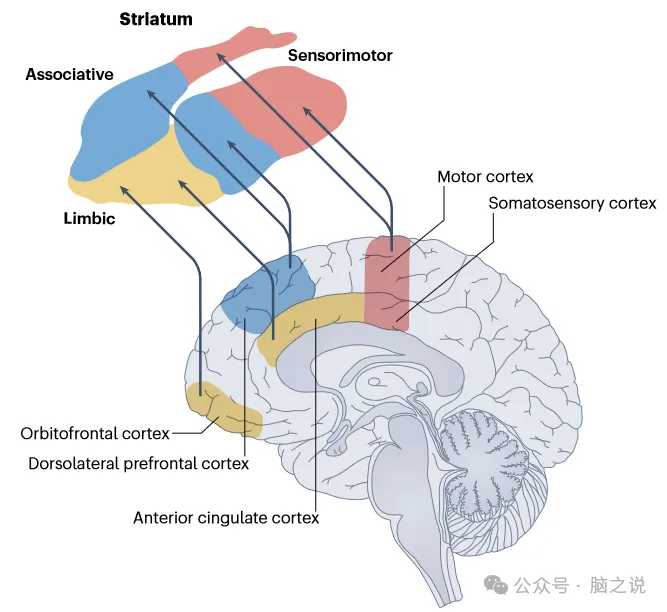
图2大脑不同区域向纹状体的投影地形图。
许多大脑区域以一种地形确定的模式投射到纹状体(图2)。一个长期存在的理论认为,精神分裂症中的多巴胺过度活跃发生在边缘区域。然而,这一假设并没有得到体内数据的支持,体内数据显示,与健康对照者相比,精神分裂症患者多巴胺合成或释放能力的最大差异在背纹状体,而不是边缘(腹侧)纹状体。精神分裂症患者与健康人相比,最大的差异是在联合纹状体区域,该区域接收额叶皮层的投射,尽管在统计上显著的升高也见于感觉运动纹状体(图2)。纹状体多巴胺合成的类似增加在其他精神疾病中也有发现,包括双相情感障碍;因此,多巴胺能功能障碍可能是一系列疾病的精神病症状的基础。
来自精神分裂症患者的死后证据和最初的体内成像显示,他们的D2/3受体水平高于神经正常个体。然而,随着大量数据的积累,最新的荟萃分析发现,相对于健康对照个体,精神分裂症患者体内D2/3受体在纹状体或皮质中没有显著的改变。此外,在一项对10名精神分裂症患者和9名健康参与者进行的PET成像研究中,没有发现D2/3受体内化改变的证据。考虑到所有抗精神病药物都是D2/3受体阻滞剂,总体D2/3受体水平缺乏重大改变令人惊讶。然而,这些测量主要反映突触后D2/3水平,突触前D2/3自身受体的改变可能存在。
多巴胺投射到皮层的密度相对较低,这使得测量这些区域的多巴胺合成或释放能力具有挑战性。然而,高亲和PET示踪剂的发展已经能够在体内测量皮质多巴胺释放能力。使用这种技术的研究发现,与健康人相比,精神分裂症患者在对安非他明作出反应时,额叶皮层和其他颅外区域的多巴胺释放量较低,在进行认知任务时额叶皮层的多巴胺释放量也较低。此外,死后的证据表明,精神分裂症患者的额叶皮质中酪氨酸羟化酶的水平比正常人低。总之,这些发现表明精神分裂症患者额叶皮层多巴胺能功能减弱。这种病理生理特征被假设为该疾病的阴性症状和认知症状的基础,尽管与症状的联系尚未得到明确证明。
3. 精神分裂症的GABA扰动
3.1 GABA的改变
氨基丁酸(GABA)是人脑中最常见的抑制性神经递质。对精神分裂症患者和正常人的死后研究主要集中在额叶皮层上。这些研究表明,与健康个体相比,精神分裂症患者的许多GABA标记物,如合成GABA和GABA膜转运蛋白的谷氨酸脱羧酶的mRNA和/或蛋白质水平较低。此外,有证据表明,在协调谷氨酸能神经元的快速放电中起关键作用的小蛋白阳性GABA能中间神经元活性较低,在患有该疾病的个体中也很明显。因此,死后证据揭示了精神分裂症患者各种GABA能标记物的改变,包括那些协调谷氨酸能神经元快速放电的标记物。
2022年对30项体内H-MRS成像研究的荟萃分析发现,与健康个体相比,首发精神病患者中扣带皮层和精神分裂症患者枕叶皮层的GABA6水平较低。然而,与健康个体相比,精神分裂症患者在其他被检查的额叶皮质区域或基底神经节中没有观察到任何变化。这些发现,再加上参与GABA产生和再摄取的蛋白质水平较低的证据,表明精神分裂症患者的皮质GABA水平较健康个体低。与首发精神病患者相比。最后,临床高危组和健康对照组的中扣带皮层没有发现差异,对此有足够的研究进行荟萃分析。
3.2 GABA受体的改变
在之前的综述中总结了多项研究,研究了使用示踪剂结合苯二氮卓类药物结合位点的GABA受体的可用性。到目前为止,这些研究还没有发现精神分裂症患者和健康人在任何被调查的大脑区域有显著差异。然而,一项研究显示,与健康个体相比,精神分裂症患者颞叶皮层中GABA受体a5亚基的示踪剂结合较低,表明该亚基在该疾病中的水平较低,尽管这一发现需要进一步的研究证实。因此,低张力抑制可能导致精神分裂症患者锥体神经元的高兴奋性。
4. 精神分裂症的基因发现
精神分裂症是高度遗传性的,大规模全基因组关联研究已经确定了200多个与精神分裂症风险相关的基因位点。虽然对这些基因座的全面回顾超出了本文的范围,但许多基因座涉及与本文讨论的神经递质相关的通路,包括D2多巴胺和mGluR3谷氨酸受体,以及与谷氨酸能传递和突触功能相关的多种蛋白质。此外,死后证据显示,精神分裂症患者大脑中多巴胺能和谷氨酸能基因的转录与健康参与者相比存在差异。此外,精神分裂症的遗传风险与尾状核中D2自身受体亚型的低表达有关。由于D2自身受体的激活减少了多巴胺的合成,因此,在精神分裂症患者中观察到,D2自身受体的低表达可能有助于纹状体多巴胺合成和释放能力的提高。
全基因组关联研究的通路分析和与精神分裂症相关的拷贝数变异研究也表明了谷氨酸能和GABAergic系统的参与。总的来说,精神分裂症的遗传风险分析与神经影像学鉴定的系统相似,表明遗传易感性导致多巴胺能、谷氨酸能和GABA能回路异常,从而导致精神分裂症。
5. 精神分裂症病理生理综合模型
最初的精神分裂症模型倾向于只关注一种神经递质。然而,在过去的十年中,影像学发现已被合并为一个综合模型,以考虑谷氨酸能、GABA能和多巴胺能系统如何相互作用,以解释该疾病的影像学发现和症状。前面章节总结的影像学结果(图3)表明精神分裂症患者神经递质改变的区域差异。这些研究通过证明多巴胺能分子改变与多巴胺合成和释放能力有关,从而完善了我们对精神分裂症中多巴胺功能障碍本质的理解,表明突触前扰动而不是转运体或突触后D2/3受体异常。研究结果还将纹状体多巴胺能最显著的改变定位于纹状体接收额叶皮层投射的部分。
成像研究表明,精神分裂症患者的纹状体和额叶皮层中多巴胺和谷氨酸的水平相反。与非精神分裂症患者相比,纹状体中这两种神经递质的水平更高,而额叶皮层中的水平更低(图3)。这些观察结果提出了GABA是否遵循类似模式的问题;需要对纹状体氨基丁酸进行更多的研究来回答这个问题。
GABA和谷氨酸的死后和体内成像结果表明,精神分裂症患者额叶皮质区兴奋性和抑制性信号发生改变,基底神经节和丘脑兴奋性信号发生改变,尽管这些改变的原因是由神经递质库还是其他机制驱动的,仍有待澄清。此外,研究结果表明,基底神经节中谷氨酸和多巴胺的改变与更严重的症状有关,尤其是在多巴胺能改变的情况下的精神病症状。

图3 精神分裂症的神经化学及其他异常。
本文的研究结果提出了一个问题,即多巴胺能、谷氨酸能和GABA能系统的异常异常是否通过共同的病理生理机制相互关联。为了支持这一理论,精神分裂症患者的纹状体多巴胺能水平较高与额叶皮层谷氨酸能水平较低相关,这种关系在非精神分裂症患者中未见。这一观察结果暗示了额纹状体回路参与了该疾病的病理生理学,尽管其潜在机制尚不清楚。一种假说认为,较低的皮质谷氨酸和NMDA受体水平导致调节谷氨酸能投射神经元的皮质GABA中间神经元的激活减少。反过来,这被认为降低了对投射到基底神经节的皮质谷氨酸能神经元的抑制,从而导致纹状体多巴胺神经元的去抑制 (图4)。因此,这种涉及从皮层到皮层下区域的投射的回路可能解释了许多与精神分裂症相关的GABA、谷氨酸和多巴胺能异常。我们把注意力集中在额叶皮质,因为它是精神分裂症患者GABA能和谷氨酸能异常改变的证据最多的大脑区域,但其他区域的异常,特别是海马体,也可能解除对皮质下回路的抑制,导致纹状体多巴胺能过度活跃。
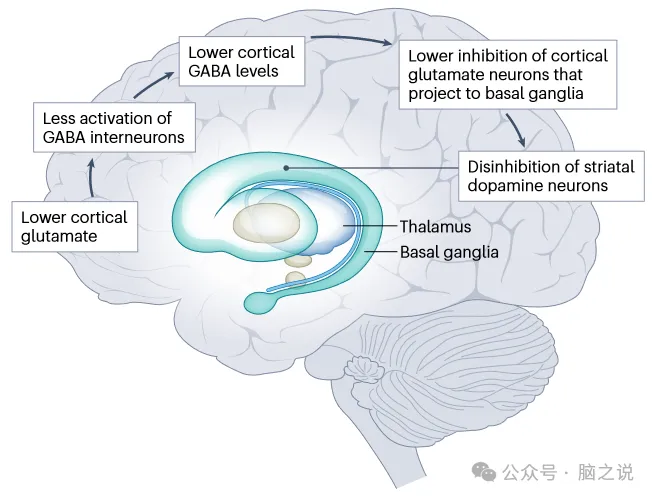
图4 精神分裂症综合模型。
5.1 潜在症状的机制
纹状体多巴胺能信号失调被认为是精神分裂症精神病症状的基础。我们对纹状体多巴胺能信号在大脑中的作用的理解取得了进展,这为这种影响是如何发生的提供了新的见解。研究表明,从中脑向纹状体投射的多巴胺神经元在预期和感知刺激之间发出信号差异,称为预测误差。中间纹状体多巴胺神经元也会调节动物的行为,使其表现得好像受到了感觉刺激,而实际上并没有——这是一种错觉。在这些发现的基础上,模型提出中纹状体多巴胺神经元的多巴胺功能障碍分别导致错误的联想和误解,从而导致妄想和幻觉(图5)。支持这一假设的研究发现,与健康个体相比,精神分裂症患者在学习任务中与预测错误信号相关的纹状体反应发生了改变。
精神分裂症认知症状和阴性症状的机制尚不明确。一种理论认为,不同的多巴胺回路参与其中,并提出认知症状是由皮层多巴胺能低下引起的,它损害了前额皮质的功能,而阴性症状是纹状体多巴胺能亢进引起的,它破坏了奖励信号。另一种理论认为,眼窝额叶皮层和前扣带皮层在自主和冷漠中起着关键作用,背侧前额叶皮层在工作记忆和执行认知功能中起着核心作用。该理论提出了这些区域内皮层微回路中兴奋性和抑制性信号的不平衡,这被假设为降低了信噪比,损害了这些区域的功能,并导致了该疾病的认知和阴性症状。
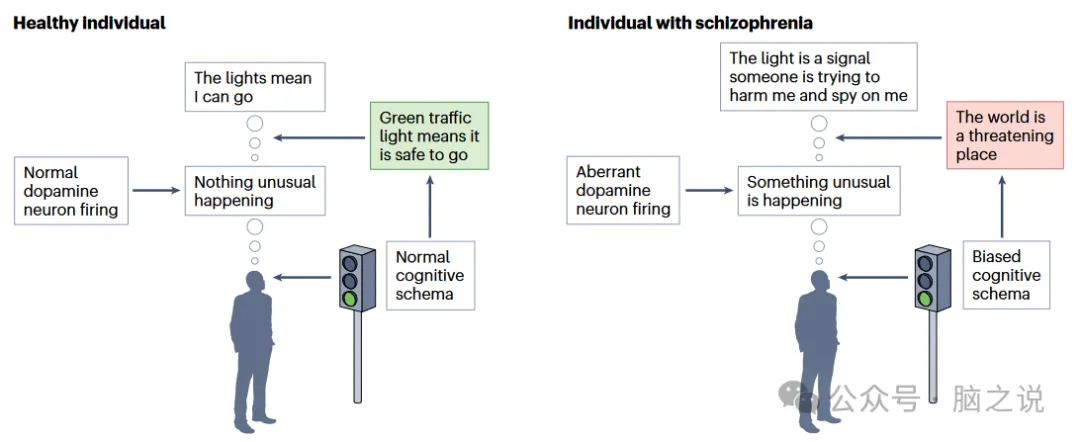
图5 多巴胺功能障碍导致错误联想和误解。
5.2 电路功能障碍的机制
综合模型所描述的电路功能障碍可以解释纹状体多巴胺合成和释放能力较高。正如“精神分裂症的遗传发现”中所讨论的,许多与精神分裂症风险密切相关的遗传位点编码参与谷氨酸能和GABA能信号通路的蛋白质。因此,精神分裂症的遗传风险可能导致大脑皮层兴奋性和抑制性信号的失衡。
精神分裂症发病的高峰年龄是从青春期晚期到成年早期。在此期间,大脑经历了神经发育的变化,额叶皮层经历了这样的过程,持续105-110年。这些变化的一个主要组成部分涉及兴奋性突触的修剪,以增加抑制活性和减少不受控制的兴奋性活动,这个过程是神经胶质细胞,尤其是小胶质细胞的关键功能。尸检和PET研究提供了精神分裂症患者额叶皮质区激活的小胶质细胞的形态学变化和标记物相对于对照组的证据。死后研究和体内PET研究也提供了证据,证明精神分裂症患者额叶区突触末端蛋白的水平低于对照组。这些发现表明精神分裂症中存在突触缺失,尽管需要进一步的研究来证实这一点。此外,在健康个体中,谷氨酸水平与同一区域的突触末端标记水平直接相关,“但在精神分裂症患者中没有发现这种关系”。这些扰动是否仅针对谷氨酸突触尚不清楚,因为迄今为止还没有研究评估体内突触变化与GABA水平的关系。然而,这些研究共同表明,精神分裂症患者的突触异常修剪改变了皮质兴奋性。
突触修剪改变可能导致皮质谷氨酸突触的丧失。如果这些突触来自刺激GABA能中间神经元的谷氨酸能侧枝,那么它们的缺失可能导致皮质抑制作用降低。反过来,正如综合模型所描述的那样,皮层下部抑制可能导致谷氨酸能投射到皮层下区域(包括纹状体)的去抑制。最后,异常的突触修剪也可能解释精神分裂症中出现的一些结构和功能成像结果,如额叶灰质体积下降和皮层活动异常。
该综合模型解释了精神分裂症中GABA能、谷氨酸能和多巴胺能异常是如何相互关联的,并通过一个涉及皮层向皮层下区域投射的回路导致症状。因此,该模型整合了精神分裂症中神经递质功能障碍的多种证据。然而,尽管这个模型为精神分裂症的神经递质改变和症状提供了一个解释,但许多假设的联系仍有待检验,而观察到的相关性并不能证明因果关系。需要进一步研究GABA能、谷氨酸能和多巴胺能之间的因果关系,并建立它们之间的时间和因果关系。此外,这里我们关注的是神经递质和神经回路,但与精神分裂症相关的大脑结构和功能的其他方面的改变也可能有助于病理生理,包括在细胞和亚细胞水平上。这些改变如何与我们所讨论的神经递质和神经回路功能障碍联系起来还有待确定,尽管研究已经开始确定潜在的机制联系。
6. 影像学结果解释的考虑
精神分裂症患者和健康个体之间多巴胺合成和释放能力改变的效应量很大;然而,谷氨酸能测量的效应量相对较低,效应量大多在0.3以下,且研究之间存在异质性。这种异质性可能与疾病持续时间和阶段、症状严重程度、某些大脑区域的低信噪比和抗精神病药物暴露等因素有关。需要进一步的工作来确认异质性是否是这些因素的结果,或者它是否表明了疾病的亚型。
H-MRS研究的荟萃分析有一些局限性。首先,个别研究的结果差异很大,分析中包括的许多研究的样本量很小,特别是那些研究GABA的研究。其次,许多研究使用了磁场低于7 T的扫描仪,与7 T的扫描相比,这与较低的测量精度有关,特别是对谷氨酰胺。因此,与较低场强相关的较低精度可能导致研究之间的异质性。最后,H-MRS不能区分细胞内和细胞外的区隔,因此精神分裂症患者基于H-MRS成像的改变可以反映一个或两个区隔的水平变化。
尽管精神分裂症患者的谷氨酸能差异明显,但这些差异是否代表体内神经传递的改变尚不清楚。谷氨酸除了作为一种神经递质之外,还具有其他功能,包括参与氮代谢和蛋白质合成,并且是GABA的前体。因此,精神分裂症的谷氨酸能差异可能与这些其他功能相关,或者与神经传递相关,或者与神经传递无关。此外,即使我们假设观察到的变化反映了突触谷氨酸和GABA,但这些变化是由突触前功能障碍、再摄取机制改变还是其他机制引起的尚不清楚。需要开发新的工具来在体内回答这些问题。
7. 对治疗发展的影响
目前所有获准用于精神分裂症的药物都是多巴胺D2/3受体阻滞剂。虽然D2/D3阻断足以治疗大多数个体的精神病症状,但并非所有个体都对治疗有反应,其他机制可能导致这些疾病。此外,D2/3受体阻滞剂对认知和阴性症状的疗效有限。此外,用这类药物治疗与锥体外不良反应有关,如静坐症、急性张力障碍、帕金森病和迟发性运动障碍,这些都与D2/3受体的阻断直接相关。其他副作用包括体重增加和代谢功能紊乱。考虑到这些限制,我们需要一种能够抑制异常多巴胺信号的新药,同时允许纹状体中的生理信号和增强皮层区域的信号。其中一种方法是瞄准D2自受体,帮助调节多巴胺神经元的功能。然而,使用这类药物的临床研究结果却喜忧参半。
7.1 TAAR1兴奋剂
目前正在开发的一种新的治疗方法针对调节多巴胺神经元放电的微量胺相关1型受体(TAAR1s)。此外,TAAR1激动剂改善了猴子和啮齿动物模型中前额叶皮层介导的与谷氨酸能功能相关的认知任务。目前有两种TAAR1激动剂处于临床试验阶段。一项使用 NCTO4512066治疗急性加重精神病患者的II期试验结果显示,与安慰剂相比,NCTO4512066在症状方面没有显著改善。
7.2 毒蕈碱受体兴奋剂
M4毒蕈碱受体作为抑制性自身受体,可减少中脑多巴胺神经元的刺激性胆碱能输入,导致下游多巴胺神经传递受到抑制。因此,使用特异性激动剂增加这些受体的传递可能会减少异常的多巴胺信号,尽管是间接的。临床前研究测试M4毒蕈碱受体的调节已经显示出治疗精神分裂症的希望。例如,xanomeline,一种M1/M4毒蕈碱受体激动剂,相对于给药的动物,用苯丙胺或MK801治疗的动物,可以减少过度运动。
在一项针对精神分裂症患者的双盲安慰剂对照先导研究中,虽然总队列规模很小,但Xanomeline治疗与安慰剂相比在症状改善方面有更大的相关性。从那时起,大规模的临床试验已经测试了这种化合物与外周毒蕈碱拮抗剂trospium联合使用,旨在阻断外周副作用。这种联合药物被命名为KarXT。KarXT在精神分裂症患者中的首次试验显示,在治疗的5周内,与安慰剂相比,阳性和阴性综合征量表和子量表的总分有了显著改善。在过去几年中,两项iii期试验取得了与初始试验相似的结果,尽管其中一项试验未能显著减少阴性症状。
其他毒蕈碱激动剂也在开发中。最先进的是emraclidine (CVL-231),一种选择性的M4毒蕈碱受体的正变构调节剂。在2022年的一项lb期试验中,与安慰剂相比,emraclidine改善了精神分裂症患者的总症状严重程度评分。该药物显示出合理的不良反应,尽管需要进一步的研究来确定其有效性。KarXT和emraclidine研究的积极结果表明,该领域可能很快就会有第一个不与多巴胺受体结合的精神分裂症的替代许可药物治疗。
7.3 治疗认知和阴性症状
鉴于D2/3阻滞剂对认知和阴性症状的疗效有限,因此需要针对这些方面的治疗。在“精神分裂症中的谷氨酸能损伤”和“精神分裂症中的GABA紊乱”中回顾的影像学证据表明,谷氨酸能和/或GABA能系统是潜在的目标。基于临床前研究显示mGLuR2/3激动剂可降低前额叶皮层谷氨酸能突触的活性,mGLuR2/3激动剂已在临床试验中进行了测试。mGluR2/3激动剂的早期临床试验发现,精神病和阴性症状均有改善,但III期试验并不成功。这些研究的一个局限性是没有测量脑谷氨酸水平,因此尚不清楚缺乏临床疗效是由于未能调节谷氨酸还是该机制与评估的症状无关。
已经开发出增强NMDA受体神经传递的药物,可能增加GABA能中间神经元的激活,从而降低投射到基底神经节的皮质谷氨酸能神经元的活性。药物作用的主要机制是增加NMDA受体协同激动剂的水平,如甘氨酸、d -丝氨酸和d -环丝氨酸,它们是NMDA受体激活所必需的。这种策略已经产生了一些有希望的试验结果,但也有许多不一致的发现,可能是由于确保足够的协同激动剂递送到大脑的挑战。为了解决这个问题,甘氨酸1型转运蛋白抑制剂已经被开发出来,目的是阻断从脑细胞释放的甘氨酸的再摄取,以增加NMDA受体的甘氨酸突触水平。其中一种药物BI425809在2021年的一项针对精神分裂症患者的双盲安慰剂对照研究中改善了认知症状。
正如“精神分裂症病理生理学综合模型”中所讨论的那样,影像学结果表明,皮层下区域的谷氨酸能和多巴胺能改变与皮层改变的方向相反。这一观察结果强调了区域选择性干预的必要性,这种干预可以促进皮层中的神经递质活动,同时减少皮层下区域的活动。脑刺激疗法,如经颅磁刺激和经颅直流电刺激,局部作用改变神经活动和功能连接。因此,这些疗法提供了一种实现区域选择性治疗的潜在途径。这类疗法在难治性抑郁症中的应用已得到证实,它们在减少精神分裂症的阳性症状、阴性症状和认知症状方面显示出一定的功效。然而,这些刺激疗法对精神分裂症的神经化学和神经生理作用还需要进一步研究。综合模型(图4)表明,抑制皮层区域突触到中脑多巴胺能神经元的谷氨酸能投射代表了另一种区域选择性方法;针对GABA中间神经元特定亚型的药物目前正在用于治疗精神分裂症。心理治疗,如认知行为疗法,在精神分裂症的治疗中也起着重要作用,并且可以通过在认知水平上起作用来补充药物治疗。
8. 精神分裂症的异质性
2017年的一项方差荟萃分析评估了精神分裂症患者灰质体积测量的可变性,揭示了与健康个体相比,精神分裂症患者的个体间可变性更大。在精神分裂症患者中,可利用性和额叶皮质区和丘脑的谷氨酸能测量也显示出更大的个体间差异。这些发现可以用该疾病的生物学亚型的存在来解释。一种说法是,对D2/3受体阻滞剂的治疗耐药性与一种特定亚型有关,其特征是谷氨酸能升高更明显,纹状体多巴胺合成和释放能力没有改变。为了支持这一假设,一些证据显示,在抗精神病治疗抵抗性或反应性精神分裂症患者的样本中,前扣带皮层谷氨酸的H-MRS成像测量呈双峰分布,与两个亚群一致;然而,本研究纳入的队列相对较小。此外,许多其他因素可能导致成像结果的异质性,包括方法学问题,如合并症的混杂效应或患者组在成像期间比对照组运动更大,从而影响数据质量。因此,需要在更大的队列中进行进一步的测试,以确认精神分裂症亚型的存在,然后才能开发精确医学方法。
另一个可能导致精神分裂症病理生理异质性的因素是先前抗精神病药物治疗的效果。对大鼠的研究表明,抗精神病药物可导致D2/3受体上调。这些结果表明,精神分裂症患者D2/3受体可用性变异性增加的潜在机制,尽管这在人类中仍有待证实。精神分裂症患者与D2/3受体阻滞剂治疗耐药相关的高未满足需求突出了理解这种变异性和探索新的潜在治疗靶点的重要性。
9. 未来方向
我们的综述强调的一个关键问题是,精神分裂症中谷氨酸和GABA的改变是神经递质水平改变的结果还是其他原因。正在开发的功能性H-MRS,用于测量谷氨酸对刺激反应的变化,为解决这个问题提供了一种方法。Jelen等人使用这种方法来评估对认知任务的反应中谷氨酸的变化。当任务增加时,健康人的前扣带皮层的谷氨酸代谢物增加——这种效应在精神分裂症患者身上没有观察到。另一项研究使用了另一种不同的认知任务,即颜色单词Stroop任务,在任务的最初部分获得了与谷氨酸相似的发现,尽管在后面的部分没有,这可能是由于随着任务的进行,学习效应对谷氨酸的影响。这些发现表明,精神分裂症患者对认知任务的谷氨酸能反应减弱,这与精神分裂症患者皮质谷氨酸能信号传导较低的理论一致(如“精神分裂症病理生理学综合模型”所述)。PET成像方法正在开发,以测量突触GABA水平的变化,以应对药理学挑战。尽管这种PET技术需要进一步验证,但一项使用这种方法的初步研究发现,与健康参与者相比,精神分裂症患者的GABA释放减弱了。
我们的综述强调,对于谷氨酸能或GABA受体亚型以及这些系统中潜在神经传递的分子机制的其他方面是否存在改变,我们知之甚少。这种知识的缺乏部分是由于缺乏适当的PET放射性示踪剂,尽管随着新示踪剂的发展,这种情况已经开始改变。例如,在出生后的正常发育过程中,NMDA受体亚单位的表达发生了转变,从主要的GluN2B表达到GluN2A表达。这一事件与突触的成熟、电路的完善和学习能力的发展相吻合。考虑到精神分裂症的发育方面,这种亚基之间的切换可能特别相关,并且只有通过测量体内GluN2亚基的变化才能完全表征。因此,GluN2B亚基的选择性放射性示踪剂将能够在精神分裂症中详细检查该亚基。鉴于越来越多的证据表明,氧化应激和免疫途径参与精神分裂症,开发针对这些途径中的元件的特异性示踪剂也将是重要的,其他未来的靶标包括突触功能和相关免疫过程的标记物,这些标记物可能参与调节精神分裂症中受影响的回路。
枕叶皮层中GABA水平较低的发现很有趣,因为该区域通常与精神分裂症的病理生理无关。相对较少的GABA研究在额叶和枕叶皮质以外的区域进行。因此,更广泛的GABA改变的存在仍然是可能的,就像多巴胺在大脑皮层的释放一样。考虑到其他证据表明颞叶皮层,特别是海马体与精神分裂症的病理生理有关,以及更严重的症状与该皮层区域低谷氨酸水平之间的关联,需要对颞叶皮层,特别是海马体进行进一步的研究。成像技术的发展使神经代谢物能够在同一个体的多个大脑区域进行测量,这将有助于解决这一问题。同样,精神分裂症病理生理中其他神经递质系统的参与仍然未知。考虑到精神分裂症患者的死后变化证据及其作为治疗靶点的潜力,5 -羟色胺能系统尤其令人感兴趣。
在这篇综述中,我们总结了神经递质因疾病阶段而异的证据,表明谷氨酸能和GABAergic65测量的改变在精神分裂症风险患者中更受解剖学限制。这些发现提供了提示进行性神经递质的重要见解。然而,这一证据主要来自横断面研究;需要额外的纵向研究来排除队列效应,并调查病理生理如何发展及其与临床结果的关系。这样的研究也可以深入了解精神分裂症的临床异质性,并有助于确定生物学亚型的存在。这些进步将使患者分层,为每个人找到最佳的治疗方法,并有助于未来的药物开发。
10. 结论
在过去的十年中,我们看到了大量关于精神分裂症病理生理学的新的体内数据。数据显示,这种疾病的特征是额叶皮质以及其他皮质区域的谷氨酸、GABA和多巴胺水平较低;基底神经节和丘脑谷氨酸水平升高;与健康个体相比,基底神经节中多巴胺的合成和释放能力更强。此外,这些改变之间的联系表明,额-丘脑-纹状体-中脑回路在该疾病的病理生理学中起着关键作用。
在皮质区域GABA和谷氨酸信号的不平衡被认为会降低神经活动的信噪比,损害皮质功能并导致认知和阴性症状。相比之下,基底神经节多巴胺能功能失调被认为会增加错误联想和误解的可能性,从而导致该疾病的精神病症状。这表明,目前的治疗方法都作用于相同的靶点,即D2受体,只解决了病理生理的一个方面。本综述中讨论的研究结果强调了新的潜在干预目标,特别是在GABA能和谷氨酸能系统中,可以解决上游改变以更有效地治疗精神分裂症。
参考文献:Schizophrenia: from neurochemistry to circuits, symptoms and treatmen.
原创声明:本文系作者授权腾讯云开发者社区发表,未经许可,不得转载。
如有侵权,请联系 cloudcommunity@tencent.com 删除。
原创声明:本文系作者授权腾讯云开发者社区发表,未经许可,不得转载。
如有侵权,请联系 cloudcommunity@tencent.com 删除。